ChIP-seq+RNA-seq如何搞定PNAS?
上个月,复旦大学生命科学学院李琳课题组与董爱武、沈文辉课题组合作在PNAS发表题为“AtINO80 represses photomorphogenesis by modulating nucleosome density and H2A.Z incorporation in light-related genes”的研究论文,揭示了染色质重塑子AtINO80参与植物光形态建成的分子机制。
光是影响植物生理和发育过程最重要的外部信号之一。在光照下,黄化(暗生长)幼苗开始光形态建成,导致下胚轴停止伸长、子叶开放和花青素/叶绿素的积累等深层形态的转变。光形态建成是连接光调控转录重编程和生物形态变化的重要发育进程。越来越多的证据表明,染色质重塑子作为转录辅助因子参与光形态和暗形态建成。然而,组蛋白变异体的掺入/排除调节光形态建成反应的机制尚不清楚。

IF=9.412
发现光形态调节因子AtINO80
RNA-seq研究AtINO80对转录组的影响
ChIP-seq研究AtINO80调控机制
进一步研究AtINO80调控的关键基因
atino80-5 和atino80-6 突变体是AtINO80(依赖ATP的染色质重塑子INOSITOL REQUIRING80 (INO80))功能缺失突变等位基因。作者比较了在红光、白光、蓝光或远红光下生长时这些突变体与野生型Col-0的表型。用显微镜测量了下胚轴伸长区的细胞长度,发现AtINO80 通过促进下胚轴细胞伸长和抑制花青素的积累而作为光形态建成的抑制因子。
光敏色素和隐花色素光受体都促进了光介导的AtINO80表达抑制。通过免疫印迹分析评估35S::AtINO80- Flag 转基因株系和pEYFP-AtINO80-G转基因幼苗中AtINO80蛋白水平。发现光照可以抑制AtINO80 mRNA的表达,但可以增加AtINO80蛋白的水平。
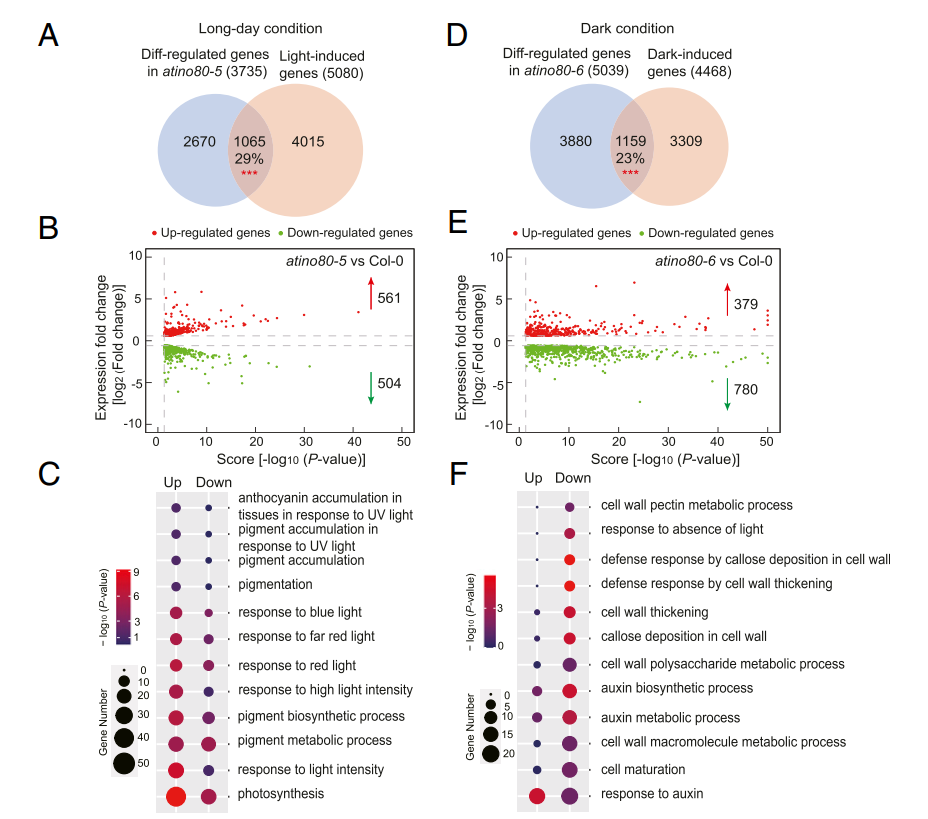
首先,通过RNA-seq比较长日照条件下生长的atino80-5 和Col-0的植株,将得到的差异基因与幼苗从黑暗转到光照条件下发现的5080个光诱导基因进行比较,发现了大量的重叠 (P < 0.001)。并对每组基因进行GO分析,表明光诱导基因,特别是AtINO80上调的基因,可能促进atino80 植物的光形态建成。
同时,作者比较了黑暗生长的atino80-6 与Col-0 幼苗,将得到DEGs与4468个暗诱导基因进行比较,发现与细胞伸长相关的基因可能有助于减少atino80 的暗形态建成。
为了研究AtINO80是如何调节转录水平,使用组蛋白H2A.Z和组蛋白H3的抗体在长日生长的Col-0和atino80-5 中进行ChIP-seq。
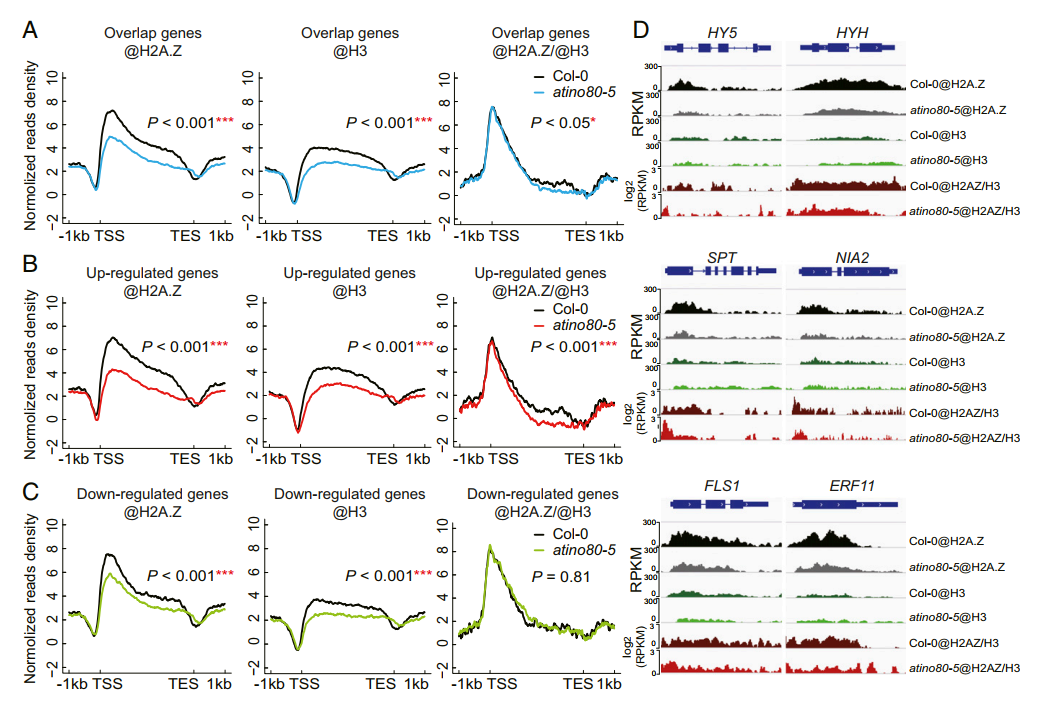
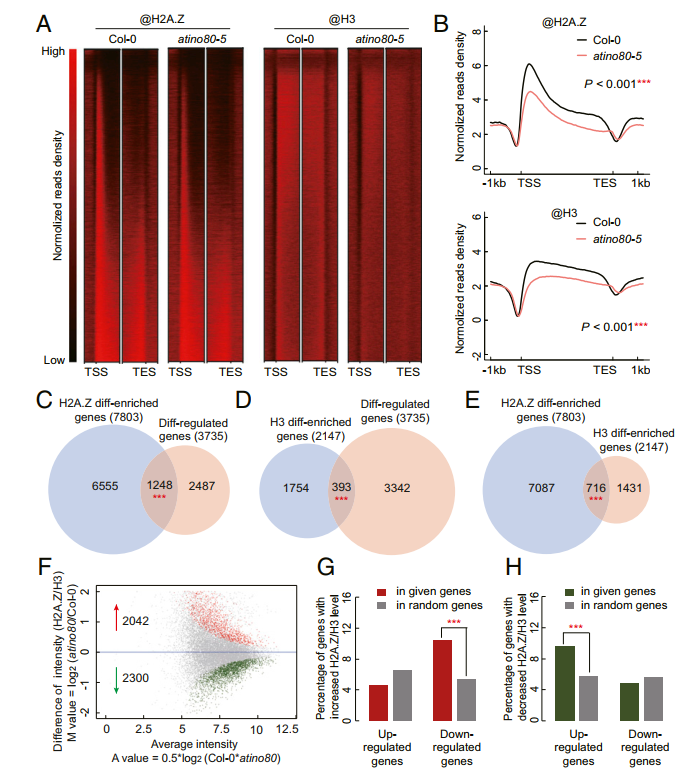
在Col-0中获得组蛋白H2A.Z约18,649个peak(19,488个基因),与已发表的组蛋白H2A.Z有很大重叠(79%)。如图2A和B所示,与前人发表的数据一致,组蛋白H2A.Z的peak分布在靠近TSS的整个gene body区。在atino80-5 突变体中,组蛋白H2A.Z水平在整个gene body区域降低。此外,组蛋白H3在atino80-5 中的占有率也发生了变化。把input作为对照,分别鉴定了7803个和2147个差异富集的组蛋白H2A.Z和H3修饰。早前在长日生长的atino80-5 中发现的DEG与组蛋白H2A和H3差异富集的基因显著重叠,表明atino80 的功能与组蛋白H2A、Z和H3的修饰水平密切相关。两组重叠基因富集在光相关的GO terms。此外,光诱导基因与组蛋白H2A.Z和组蛋白H3差异富集基因有显著重叠。
组蛋白H2A.Z标记可能受组蛋白H3占有率变化的影响,因为组蛋白H3差异富集基因和组蛋白H2A.Z差异富集基因之间存在大量重叠(P<0.001)。为此,把组蛋白H3作为input,对组蛋白H2A.Z进行差异分析,对所有组蛋白H2A.Z富集区域进行差异富集分析。在atino80-5 中,鉴定到了2300个组蛋白H2A.Z减少的区域和2042个组蛋白H2A.Z增加的区域。atino80-5 中上调的DEGs在组蛋白H2A.Z降低的基因中显著过表达,而下调的DEGs在组蛋白H2A.Z升高的基因中显著过表达。表明组蛋白H2A.Z的占有率与AtINO80调控基因的转录呈负相关,这与最近的报道一致。
接下来,作者将重点放在光响应基因上,以确定组蛋白H2A.Z、H3的变化程度,以及组蛋白H2A.Z/H3在光和AtINO80转录调控中的比例。观察到重叠基因中组蛋白H2A.Z/H3减少(P<0.05,),与Col-0相比,atino80-5 中上调的基因更为明显(P<0.001)。然而,在光和AtINO80调控基因重叠基因群中,组蛋白H2A.Z/H3修饰的降低并不显著(P =0.81)。
作者追踪了特定基因中H2A.Z和H3组蛋白的水平。组蛋白H2A.Z和H3在HY5、HYH、SPT、NIA2、FLS1 和EFR11 gene body富集减少可能导致它们在atino80 中的转录水平升高。值得注意的是,AtINO80增加了组蛋白H2A.Z、H3和H2A.Z/H3在HY5 gene body的富集,从而抑制了HY5 的表达。HY5 是幼苗光形态建成过程中的一个关键调控因子,调控多种基因的转录。因此,AtINO80对上述基因表达的影响可能是通过改变组蛋白H2A.Z水平和核小体密度来控制HY5 的转录的结果。
最近发表的一篇论文表明,HY5 的主要活性是通过直接靶向127个基因来促进转录的。在长日生长的atino80-5 中鉴定出的DEGs与这些直接的HY5 靶基因有显著的重叠(P<0.001)。AtINO80对这些重叠的HY5靶基因表达的作用可能是由于组蛋白H2A.Z和H3水平的降低,比如RGA-LIKE3(RGL3),microRNA 163(miR163), RESPONSIVE TODESICCATION26(RD26), At1g25400和CYS, MET, PRO,AND GLY PROTEIN1(CMPG1),也可能是HY5 水平增加的结果,比如对于At4g32480, BTB AND TAZ DOMAIN PROTEIN 5(BT5), MYB44, At4g38825和REDUCED EPIDERMAL FLUORESCENCE5(REF5) 基因。
综上所述,AtINO80在光调节基因表达中的作用可能是通过改变核小体密度和组蛋白H2A.Z水平直接调控的,也可能是通过其下游基因HY5 间接调控的。