
R-loop CUT&Tag系列文章之二:R-loop与表观调控
承接上篇R-loop概览,本篇文章我们将介绍R-loop与表观遗传学调控的关系,主要针对染色质可及性和转录调节。
R-loop相关的染色质特征(组蛋白翻译后修饰)与启动子和转录起始位点处的转录特征类似,包括组蛋白H3 Lys4的单甲基化和三甲基化(H3K4me1和H3K4me3)、组蛋白H3 Lys27乙酰化(H3K27ac)和转录延伸(组蛋白H3 Lys36的三甲基化,H3K36me3)。R-loop可以诱导染色质去凝聚(chromatin decondensation),也可以促进异染色质组装(heterochromatin assembly)和染色质压实(chromatin compaction)。

在RASSF1A位点,反义长链非编码RNA(lncRNA)RASSF1反义RNA 1(ANRASSF1)在启动子区域形成R-loop,可招募PRC2(polycomb repressive complex 2),通过H3K27me3来沉默RASSF1A表达。
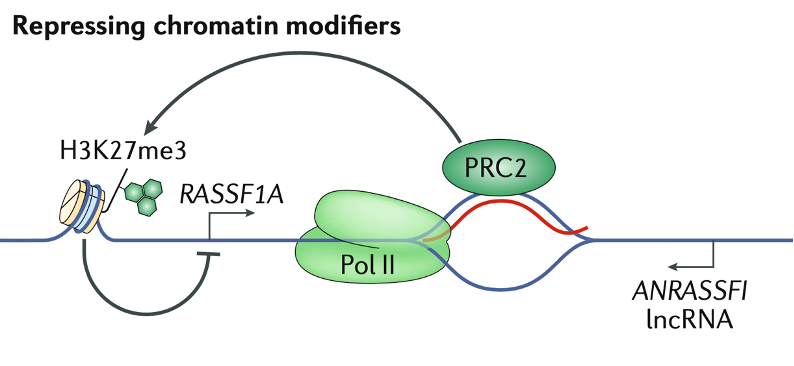
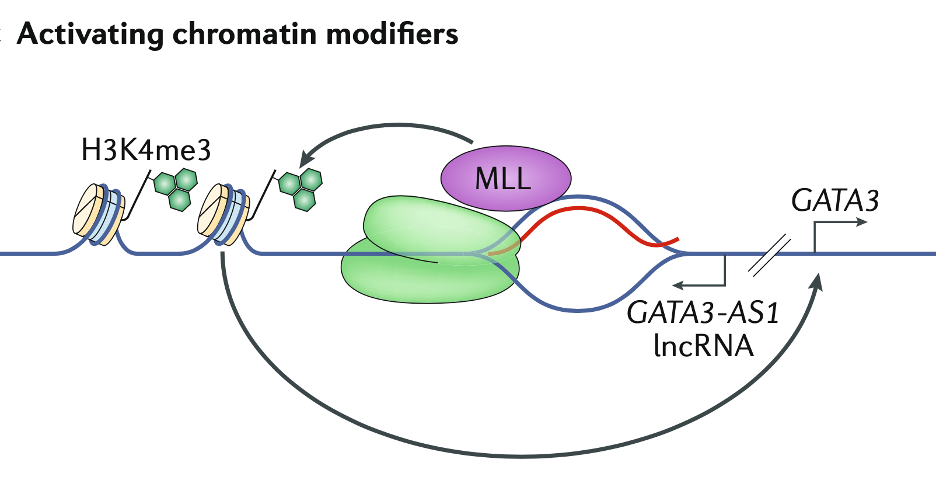
染色质修饰剂也可促进转录。例如甲基转移酶复合物MLL(mixed-lineage leukaemia)家族,被GATA3基因启动子区域的R-loop招募,通过H3K4me3促进转录。
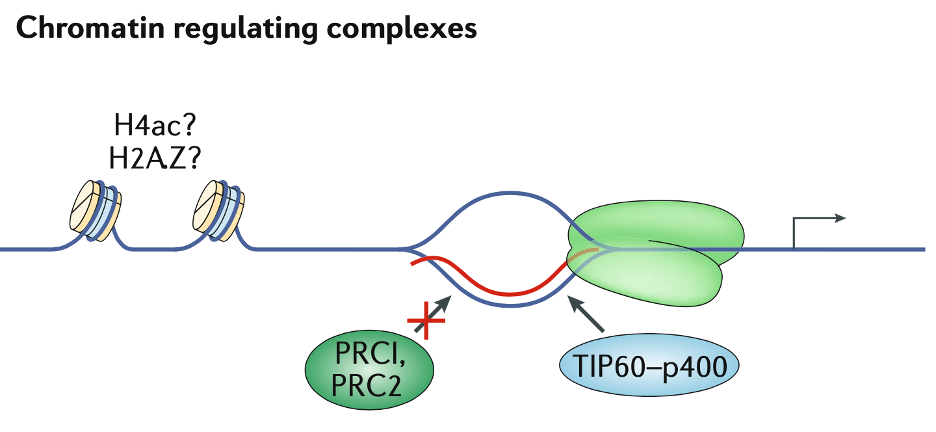
在胚胎干细胞分化过程中,R-loop形成趋势可以决定选择哪种类型的染色质修饰剂结合。例如具有强烈R环形成趋势的启动子区域不结合抑制型PRC2或PRC;相反,其募集TIP60-p400并激活转录。这可能引起组蛋白修饰,例如组蛋白H4乙酰化 (H4ac)或组蛋白变体(H2A.Z)。
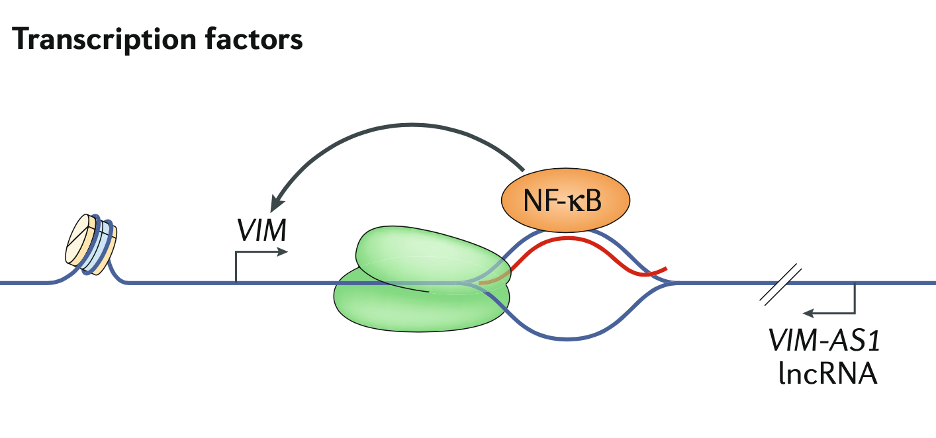
R-loop可以招募转录因子来促进基因表达。在VIM位点,反义lncRNA VIM的反义RNA 1(VIM-AS1)通过在转录起始位点形成R-loop来促进NF-κB(nuclear factor-κB)募集。
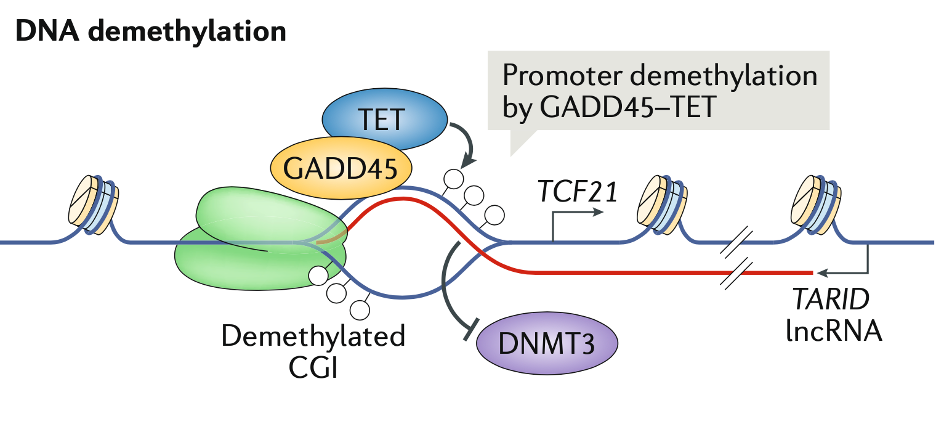
在部分含有 CpG 岛 (CGI)的启动子中,GADD45(growth arrest and DNA damage protein 45A)充当 R 环“reader”,招募DNA去甲基化酶TET(ten–eleven translocation),使启动子 DNA 去甲基化,激活转录。下图显示的是lncRNA TARID(TCF21 antisense RNA inducing demethylation)和顺式靶基因转录因子TCF21(gene transcription factor 21)。从富含R-loop的CGI去除DNMTs(DNA (cytosine-5)-methyltransferases),例如DNMT3,可能可以增强去甲基化的效果。

上述调节性R-loop与染色质可及性和转录调节总结如下表:
序号 | R-loop中RNA | R-LOOP位置 | 功能及影响 |
1 | ANRASSF1A | RASSF1A启动子 | H3K27me3沉积和RASSF1A抑制 |
2 | GATA3-AS1 | GATA3启动子 | H3K4me3沉积和GATA3转录 |
3 | TIP60–p400依赖基因 | 胚胎干细胞中lncRNA亚群 | 防止PRC介导的基因抑制 |
4 | VIM-AS1 | VIM启动子 | NF-κB募集与VIM1转录 |
5 | TARID | TCF21启动子 | DNA去甲基化和TCF21抑制 |

