
国内首篇! 一区IF 13.584 I 嘉因生物CUT&Tag助力大刺鳅性染色体起源的研究
客户寄语:
CUT&Tag是研究蛋白质与DNA互作的新方法。与传统ChIP-Seq相比,该技术操作简单,信噪比高。我们也是**次尝试用CUT&Tag来做H3K9me3修饰的ChIP,效果还蛮好的,对文章的提升帮助很大。公司的技术人员也积极向我们解释这个新技术的原理以及数据的处理的细节,合作比较愉快。
性染色体的起源需要在原性染色体之间建立重组抑制。在许多鱼类中,性染色体对是最近起源的同态染色体,为研究重组抑制如何以及为什么在性染色体分化的初始阶段进化提供了物种,但这需要高完整度的基因组。
2021年7月,福建淡水水产研究所发表“Telomere-to-telomere assembly of a fish Y chromosome reveals the origin of a young sex chromosome pair”一文,首次发现大刺鳅(Mastacembelus armatus)间性群体,组装出完整度极高的基因组,提出动物性染色体近着丝粒起源的假说,并对其重组抑制进行了相关研究。

基于已知但染色体水平装配不完整的大刺鳅基因组(fMasArm1.2),结合本文三代测序(PacBio RS II)获取的约30G的高保真序列(HiFi)数据,组装获得大刺鳅的基因组序列,其可独立产生两个染色体水平的单倍体基因组hap-X和hap-Y。其中hap-Y基因组中24条染色体的gap数均小于10,平均gap数为5.5,而hap-X基因组的平均gap为6.4,故在以下分析中,除非另有说明,否则使用hap-Y基因组。
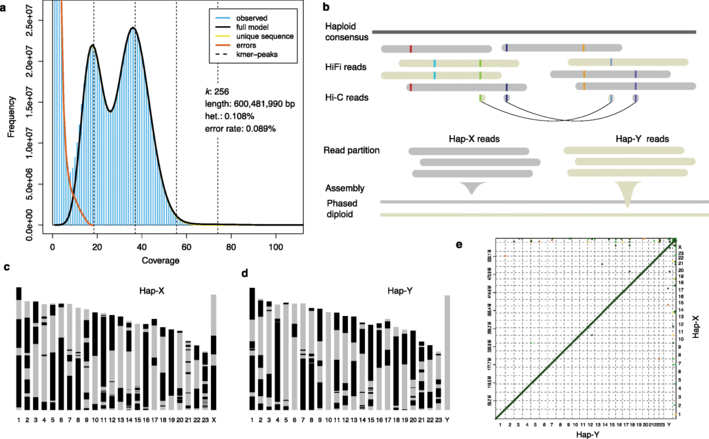
图1 单倍型基因组的组装
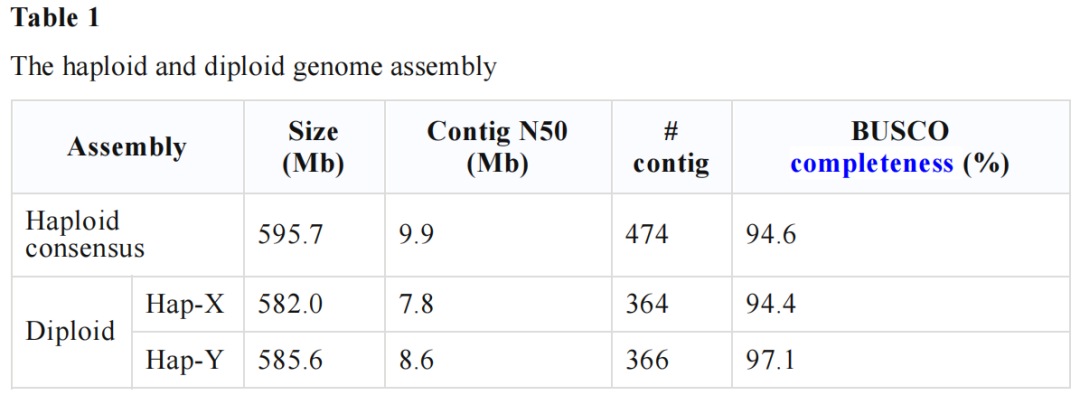
根据细胞遗传学观察大刺鳅二倍体的48(2n)条染色体,将其分为5对中着丝粒染色体、3对亚中着丝粒染色体和16对端着丝粒染色体。着丝粒结构复杂,含有大阵列串联重复序列,利用上述高质量基因组获取两个卫星序,单体长度分别为524(CEN-524)和190bp(Tel-190)。CEN-524通常出现在每个染色体上的单个位点上,在端着丝粒染色体中,它出现在一个染色体末端,而在中着丝粒和亚着丝粒染色体中出现在中间。这使得Cen-524成为着丝粒卫星的有力候选。而Tel-190仅出现在染色体末端,在中着丝粒染色体上有时出现在两端,表明Tel-190与端粒有关。为了进一步验证Cen-524为候选着丝粒卫星,利用荧光原位杂交(FISH)对Cen-524和Tel-190探针进行杂交,发现它们在染色体上的位置与基因组序列组装基本一致。
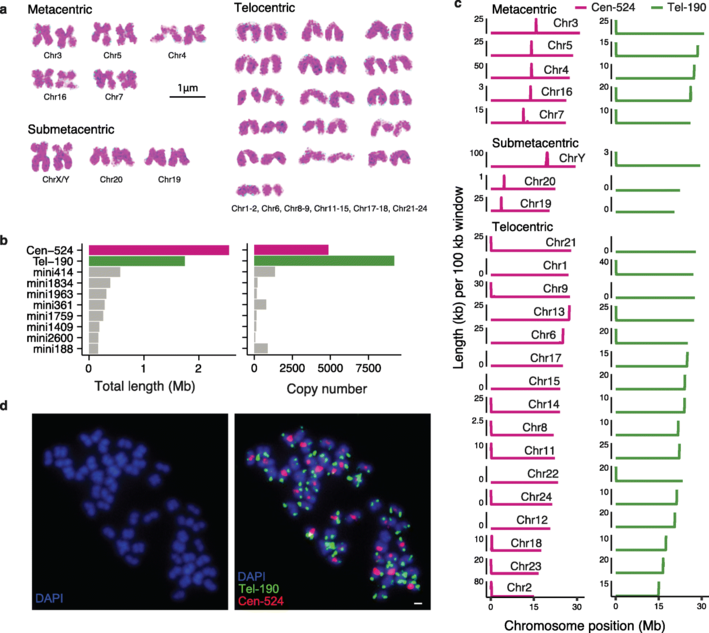
图2 核型和着丝粒的鉴定
对雌雄各10例个体进行重测序,并对与性别相关的变异进行筛选,发现Y染色体上的~7 Mb性连锁区(sex-linked region,SLR)序列,与假常染色体区(PAR)或常染色体相比,显示出高密度的雄性特异性突变。据此推测SLR跨越着丝粒,染色体的两端是PAR。这表明,物理上靠近着丝粒的位置可能是SLR缺乏重组的原因。根据雄性特异性突变的密度和内含子序列的X-Y差异,SLR可分为R1和R2两个区域。R1长约3.0Mb,包含67个蛋白质编码基因并跨越着丝粒,而R2(3.9Mb)仅包含22个基因,并且R1较R2有更高的雄性特异性SNP密度。在R1和R2中,X-Y序列差异接近1%,表明是最近的起源,我们没有检测到定义这两个区域的染色体倒置(inversions)或X、Y染色体间基因含量的差异。
大部分SLR在X和Y染色体上都有高密度的重复序列,性染色体对的整个短臂,包括X和Y染色体的着丝粒区域,具有异染色质的典型特征,包含大量的H3K9me3修饰(嘉因生物利用CUT&Tag技术获得基因组上H3K9me3修饰信息)。由于短臂异染色质位于着丝粒附近,我们推测短臂异染色质可能起源于着丝粒周围异染色质(PCH)。
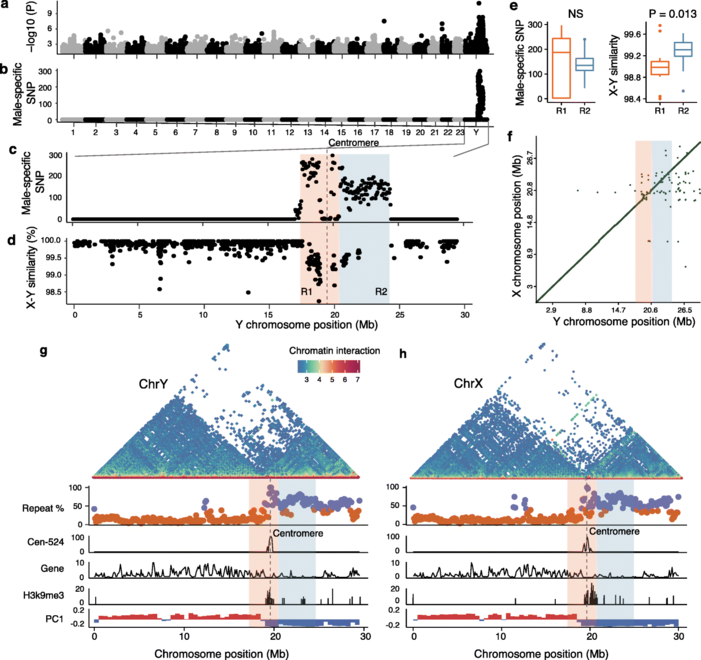
图3 性连锁区域的鉴定
检测雄性、雌性以及中间性个体的性腺中SLR基因的表达谱,获得两个特异性表达的基因syce3和hmgn6。syce3只在睾丸和卵睾体(ovotestis)后期中表达,表明其可能参与成熟睾丸的精子发生或其他生物学过程,而hmgn6在睾丸和卵睾体(ovotestis)早期表达水平相似,但在卵巢(ovaries)中的表达水平要低得多,这使其成为指导睾丸发育的性别决定候选基因。hmgn6有一个HMG(high mobility group)结构域,该结构域也存在于哺乳动物的主性别决定基因Sry中。syce3和hmgn6都位于R1,syce3非常接近SLR和PAR的边界。
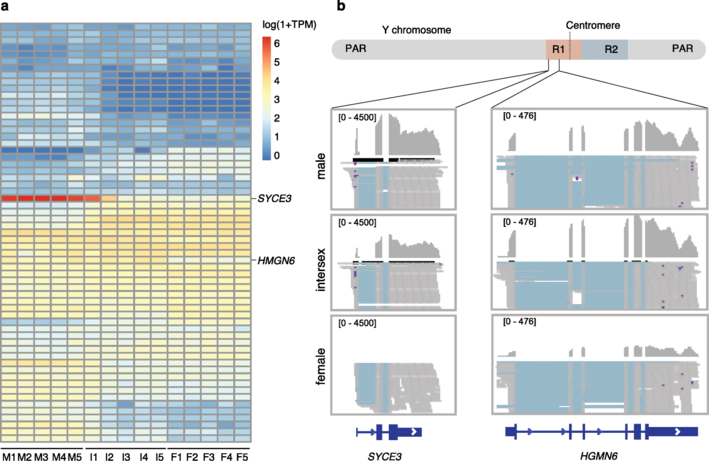
图4 HMGN6和SYCE3在生殖腺中的表达
接下来探讨常染色体是否也有类似在XY性染色体中所见的大的着丝粒周围区域。结果显示着丝粒周围的区域(~4 Mb)具有较高的重复序列(通常高于50%)、较低的基因密度、较低的重组率和更频繁的H3k9me3修饰,与PCH一致。大多数PCH长约4.2 Mb,其大小仅与染色体大小呈弱相关且不显著,因此较小的染色体,特别是端着丝粒和近中着丝粒染色体,具有较大比例的PCH,包括XY染色体。在着丝粒周围区域,物理距离较远的染色质相互作用更为频繁,符合其较高的折叠和压缩程度特征。但出乎意料的是着丝粒周围区域比其他区域有更大比例的高表达水平和宽度的基因,这与先前的观点一致,即H3K9me3在基因抑制中的作用有限,并且可能存在其他表观遗传修饰调节PCH中的基因表达。
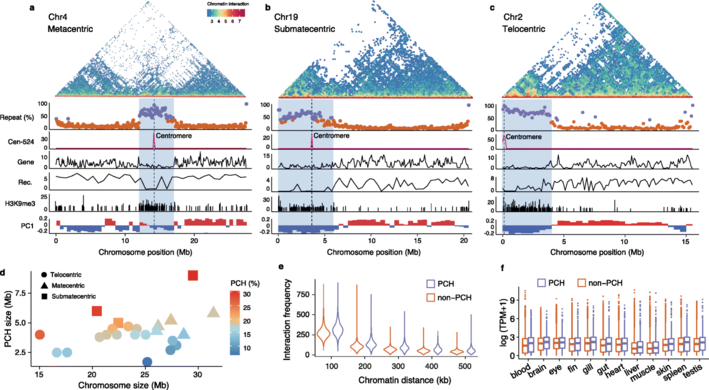
图5 着丝粒间异染色质对基因组区段化的影响
本文结合三代测序和HiFi数据从染色体层面上有效地组装大刺鳅单倍型基因组。该基因组包含大多数染色体的亚端粒和着丝粒周围异染色质序列,包括X和Y染色体。SLR位于着丝粒周围区域,表明祖先的低重组区域可以产生SLR而不依赖于对重组抑制的自然选择。